In a previous article, some of the ways in which creationism distorts and attacks a caricature of evolution were exposed, by discussing the topic of chance in evolution: creationists claim that evolution supposedly says that life emerged out of mere chance. In this article, another creationist distortion of evolution will be examined; namely, the attack of creationism on the fossil record and its gaps. Please note that the previous article is a prerequisite for understanding properly the present one.
According to creationists, the gaps in the fossil record — which are real — do not show a gradual and smooth change of one species changing into another one, but show what creationism would “predict”, assuming a special creation by God who creates species at some point in time: the fossil record shows species changing abruptly.
| But
precisely! Modern evolutionary theory claims that species do not change gradually, but “abruptly”! |
The idea that species “morph” gradually into other species is an old one, dating to Charles Darwin’s time (mid-19th C.), though not necessarily to Darwin himself.(*) Back then evidence was scarce, so it was impossible to determine the rates at which species change. Moreover, there were some antagonistic theories, such as Lamarck’s,(*) which claimed that individuals (not just species) could acquire novel traits from the environment, and pass them to their offspring. For example, according to Lamarckism, the iron smith acquires strong muscles by working daily with the iron, and that’s why his children are also strong. Also, Georges Cuvier had supported the theory of catastrophism, a consequence of which concluded that species appear relatively suddenly after catastrophic geological events.(*) It was against this background of a mixture of misunderstanding and confusion that Darwinists claimed that species (not individuals) change gradually by means of natural selection, over the course of thousands or millions of years. Many biologists who argued for evolution after Darwin’s time (early 20th C.) assumed a wrong model, according to which species have some supposed innate tendency to evolve; thus, if nothing happens, it was thought that species will gradually evolve to different species.
This idea, although in line with the Victorian prevailing thought of inherent progress in society, was nonetheless wrong. As was mentioned in the previous article, species “reflect” (in an abstract way) the properties of their environment. If nothing that threatens the survival of a species happens in its environment, the species will live “happily” in its stable niche. But if — as we saw in the example of the aquatic frogs that turned to tree-dwellers — an environmental pressure appears that threatens its survival, then the originally stable and “happy” species might give rise to a number of short-lived transitional species, until one or more of the transitional species find new stable niches (if that happens), and live there “happily” evereafter, seen not as transitional anymore but as stable,(*) until the next environmental pressure, of course. The short-lived transitional species, together with some other reasons that will be explained soon, are the cause of the observed gaps in the fossil record.
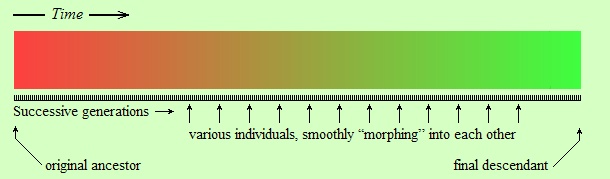
Let’s make a drawing of this idea. First, the idea of gradual constant change(*) is shown schematically in the following figure:
The original ancestor species is represented by individuals of red color in the above figure. Each individual gives birth to another individual, represented by the next little black line under the colored strip. Each descendant differs from its parent (in color in this figure, but in physical traits in reality), but the difference is so tiny that we cannot notice it merely by looking at the parent and the child. However, looking at a sufficiently distant descendant (green color) over a long period of time we can see that there is a definite difference (red vs. green) between original ancestor and distant descendant.
But, as I said, this view is most likely wrong. The following figure seems to be a more accurate representation of reality:
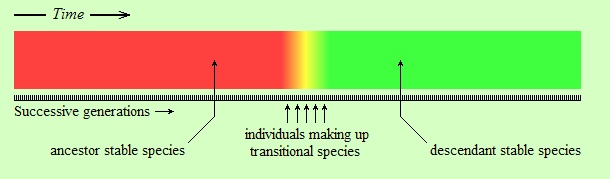
The difference in the above figure is that the change is relatively abrupt. The word “relatively” in this context is important. The individuals shown in orange, yellow, and lime in the figure (labeled as “individuals making up transitional species”) might have lived for several thousand years. However, the entire colored strip from left (red) to right (green) is supposed to have lasted for tens of millions of years, or more. Thus, the transitional species are represented by a very narrow piece of time on the strip. Therein lies the first reason of the fossil record appearing as having gaps. Because, suppose that you can take a few random snapshots within this time interval, represented by the black vertical lines on the following strip (assume each black line is a fossil):
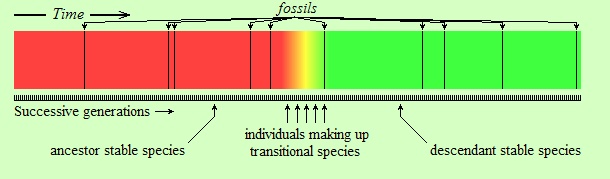
As you see, the snapshots (fossils), being random and sparse, “missed” the transitional species (orange, yellow, and lime) entirely. In reality, the reader should imagine a much narrower interval of time for transitional species than the one shown above, and a sparser set of fossil snapshots. Consider, if the length of the entire strip is, say, 10 million years, and if the length of the transitional period is 50 thousand years (reasonable numbers for evolutionary standards, depending on the kind of living beings considered),(*) then the transitional period would be only five thousandths of the entire length; it would take a width of only about two pixels to represent it in the above figure. Imagine having to “hit” that kind of interval in your sample of fossils. It is in this sense that we say that species evolve to other species “abruptly”: not literally abruptly (not from parent to child, of course, as Lamarck thought), but so fast that when seen from a faraway perspective the transitional period almost vanishes, and the change appears as if it is abrupt.
The above model of abrupt evolutionary change is called the “punctuated equilibrium” model. It says that long periods of little change (relative “equilibrium”) are interrupted (“punctuated”) by short periods of rapid change.(*)
In addition to the above, there is one more reason that contributes to the scarcity of transitional fossils:
| Fossilization is an extremely rare event |
Just because a creature dies does not imply it will become a fossil! Special conditions must be present for fossilization to take place. For example, favorable conditions include the dead creature being covered by the volcanic ash of a nearby volcano that erupted, or by the mud of a lake or river that flooded. But not all creatures live close to an erupting volcano, or by the banks of a river! Even then, such conditions do not guarantee the preservation of the creature as a fossil; they only increase the probabilities that fossilization will take place. Lastly, a creature must contain some hard parts, such as bones, to fossilize; bacteria, jellies, worms, and other soft-bodied creatures, generally do not leave fossils behind. According to Richard Cowen (in History of Life) the majority of animal species living now are unlikely ever to leave fossils, since they are soft-bodied invertebrates such as worms and slugs. Consider that of the more than 30 phyla of living animals, two-thirds of these have never been found as fossils! For all the above reasons, transition from “red” to “green” might happen, as in the previous figures, but if none of the “yellow”, etc., transitional individuals fossilized, then the change went undocumented in the fossil record. No matter how hard we dig, a transitional individual will not be unearthed, because none is there to be found. In that case it is not the paleontologist’s ax, but nature itself that “missed” recording the transition.
And yet, the savvy reader would think, there isn’t only a single instance in which a species evolved to something else; there have been billions of instances of evolution. Missed once, missed twice — well, at some point we should discover a case of transitional fossils. Isn’t that only logical?
Exactly, it is. And that’s precisely what has occurred: in a few lucky cases — bingo! Paleontologists unearthed transitional fossils.
Now, what happens in such cases, as a rule, is that creationists, donning the spectacles of experts — which typically they’re not — deny that these are transitional species. (They deny even more incredible things, such as the scientific methods used in dating fossils; that is, they deny elementary particle physics; so this denial is not too surprising.) So, if an “orange” fossil is unearthed, they’ll say, “Oh, this is really red!”; if it’s a “lime” fossil, then they’ll go — guess — “Oh, come on, that’s green folks! Don’t you see it?”; and if it’s a “yellow”, then I suppose they toss a coin and determine to call the fossil either “red” or “green”, whatever makes more sense to them.
However, unfortunately for them, reality is ruthless. Sometimes it has happened that the transition occurred between so vastly different life-forms that the transitional species that emerged cannot possibly be lumped into either the red, or the green category, to the creationists’ chagrin. Such was the situation when aquatic vertebrates (bony fish), in the course of millions of years, “came out” of the water and “conquered” the land, first yielding literally “fish with legs”, i.e., transitional creatures that disappeared (but will be presented below), then the amphibians, and from them the reptiles, mammals, and birds. Here, “came out of the water” does not mean that a single individual fish acquired legs and started wandering triumphantly on land! It means that, species after species, along some particular evolutionary branch within the tree of evolution, individuals were becoming more adapted to life on land, and less adapted to living in water. Of course, that evolution resembles a tree (rather than a linear chain) means that at the same time other branches of the tree were evolving but not toward moving out of water. The transition that yielded amphibians from fish is very important from an evolutionary perspective, not only because have transitional creatures been found and will be shown below, but also because by studying this transition we learn multiple important principles of evolution. Specifically,
Hmm... did I say “frogs”? Why don’t we let our friend, the tree frog, who explained to us the story of its ancestors in the previous article, to come back and tell us how its much-much earlier ancestors, the fish, came out of the water. Note that although the story will be recounted by a froggy, it’s not “just a story”, or a legend, but an account of that particular phase of vertebrate evolution that has been amply documented with fossil evidence. Let’s give the podium to the frog now.
 |
Hello again! Remember me? Yup, same chap, the one who told you the legendary story of how our ancestors took to the trees, yielding us. This time I’ll recount not a legend, but reality, and will show you the evidence for what I’ll say — wait and see. I’ll tell you how the land was conquered; or rather, re-conquered. You see, we have to go back to around 400 million years ago, and in those times there were fish in the water, but on land there were only plants and non-vertebrate animals, such as insects, which had already conquered the land at a much earlier time. Those insects and plants became the untapped food resources of the new explorers of land that were about to appear. |
Let’s take a look at life in the water back then. There were already plenty of fish, which, up to our days, are divided into bony fish, which includes most of our familiar ones, and cartilaginous fish, which means: sharks, rays, and chimeras. These cartilaginous fish are the more ancient, more “primitive” ones, because they appeared first. They don’t have bones, but cartilage, the white rubbery substance that you see between bone and muscle tissue when you eat chicken. (Yuck! how can you people eat chicken!) Bone seems to be a “better idea” than cartilage from an evolutionary standpoint, because when bony fish appeared they fared better in survival terms and displaced the once thriving cartilaginous fish. That’s why today the latter are restricted to only a few specialized niches, occupied by sharks, rays, and chimeras.
Bony fish are again subdivided into two large categories: the lobe-finned fish, and the ray-finned fish. As before (with the cartilaginous vs. bony distinction), today the two groups are not equal in size. There are only eight surviving species of lobe-finned bony fish in the world (2 coelacanths, 1 Australian lungfish, and 5 South American and African lungfish). Compare that to more than 22,000 species of ray-finned bony fish! Practically all of your familiar fish are ray-finned; they are the “winners” of aquatic vertebrate body plans in evolution.
 |
 |
|
| A lobe-finned fish: the Australian lungfish (Neoceratodus forsteri) © (*) | A typical ray-finned fish: the kelp bass (Paralabrax clathratus) © (*) |
And yet, here we’re concerned with the tiny group of lobe-finned fish, which was much more diverse back then, because it is from them that our common ancestors, the amphibians, emerged. According to this Wikipedia article, lobe-finned fish “are bony fish with paired rounded fins. These fins, being similar to limbs, suggest that these fish may be ancestors of land vertebrates.”
Among the lobe-finned bony fish that existed back then, there were two main types: the coelacanths, which lived in the oceans (and, as I said above, only two species of them survive today), and the rhipidistians, which are of particular interest to us. (See the skeletons of two rhipidistian fossils below.) The rhipidistian fishes split again into two main groups: the lungfish (one surviving descendant of which is pictured above), and the tetrapodomorphs. Guess what: we are all descendants of the tetrapodomorphs.
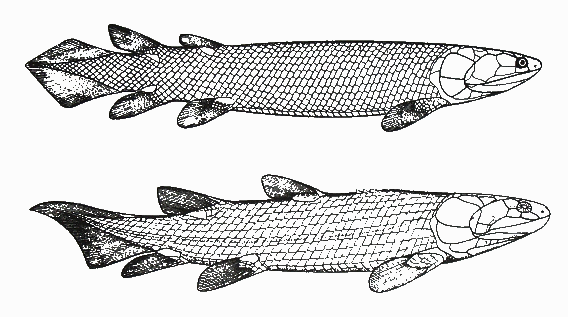
Drawings of fossils of rhipidistian fish: Gyroptychius (up) and Osteolepis (down) (from R. Cowen’s History of Life, p. 153)
Did the prefixes “lung-” in lungfish, and “tetra-” (four-) in tetrapodomorphs ring any bell? That’s right, the lungfish developed primitive lungs, by which they can breathe air, so they have two ways of acquiring oxygen: through the air and through water, because they also have gills like all other fish. They need their lungs because they live in freshwater habitats that can become muddy because of desiccation, so the oxygen content in such waters is very low, and the lungfish then use their lungs to breathe. This doesn’t mean that our lungs are direct descendants of the lungfish lungs, because we said our ancestors came from the other group, the tetrapodomorphs, which is Greek for “four-feet-like” creatures (their four underside fins were strong and good for letting them “walk” on the bottom of their freshwater habitats). Many of them developed lungs like the lungfish, because they also lived in shallow, muddy places.
Now, just how did lungs evolve in fish? One possible way is by initially swallowing a bubble of air. Many fish exhibit this behavior today, even gobies and goldfish: they come to the surface, open their mouths, and “bite” a bubble of air. They keep the bubble in their mouth for awhile, until the oxygen is diffused in their blood and the bubble shrinks, containing mostly CO2, at which point they spit it out. You’ve probably seen them in aquaria, doing this thing. Fish that live in waters with not very high concentration in oxygen can use this alternative method of breathing. Tetrapodomorphs probably developed lungs because of the habit of biting bubbles of air. Of course, when we say “developled lungs” we don’t mean that a single fish developed lungs! We mean that generation after generation, for thousands of generations, the primitive mechanism of absorbing the oxygen from the air bubble was perfected, and the organ that we call a “lung” gradually took shape and became more efficient. But, remembering the colored strips at the beginning of this page, what is “gradual” when seen from up close is “abrupt” when seen from faraway.
Likewise, tetrapodomorphs perfected their ability to use their limbs in order to slither for awhile on land, sometimes half submerged in the water and half out of it. Why did they do that? There are various reasons that have been suggested: they could be using the sunlight for basking, like modern reptiles do; or they could be moving on land to nearby ponds to spawn their eggs there. Whatever the reason, in a few million years the tetrapodomorph rhipidistian fish acquired features that made them look as if they were mixtures of fish and early amphibians. One such in-between creature, the Tiktaalik, is shown in the drawing, below.

Tiktaalik roseae, an early tetrapodomorph (late Devonian period, ~380 M. y. ago) (Credit: Arthur Weasley, GNU Free Documentation licence)
Of course, the colors shown in the above figure are there by the artist’s “poetic license”. The shape of the body, however, the leg-like fins, the amphibian-like head, etc., are all accurate renditions derived from fossil skeletons, which were found on Ellesmere island, Nunavut, in northern Canada, and date back to approximately 380 million years ago. At that time, that part of today’s Canadian land was close to the equator. Compare Tiktaalik also with the earlier picture of the lungfish, and notice the similarities in the shape of the tail, the location of the fins-legs, and the flattened shape of the head. But one essential difference is that the Tiktaalik shows that tetrapodomorphs were on their way to developing a neck, i.e., the ability to move the head independently of the body, a feature missing from the lungfish but essential for any creature that raises its head out of the water, and doesn’t always swim like a fish.
The Tiktaalik is a transitional creature — as transitional as it gets. To ask for something more transitional than that (dear creationists), is pathetic; it implies you don’t want to believe your eyes. According to this article in Wikipedia, the Tiktaalik had the following blend of features:
But of course the Tiktaalik is not the only intermediate, transitional fossil ever found; it is just one of the most recent ones. (The article announcing its discovery was published in Nature in April 6, 2006.) Since decades ago other transitional fossils between fish and amphibians have paraded in biology textbooks, such as the Ichthyostega, the Acanthostega, and others. Those are creatures that lived later, between 365 and 350 million years ago.

Ichthyostega, a late Devonian (~365 M. y. ago) transitional tetrapodomorph (Credit: Arthur Weasley, GNU Free Documentation licence)
Notice the more developed webbed feet of the Ichthyostega (meaning “fish-roof”), above, and compare them with those of the Tiktaalik.

Acanthostega gunnari, another late Devonian (~365 M. y. ago) transitional tetrapodomorph (Credit: Arthur Weasley, GNU Free Documentation licence)
Acanthostega (meaning “thorn-roof”) also had webbed feet like Ichthyostega, but was more adapted to aquatic life. This doesn’t mean that evolution moved “backward”! At any point in time after the out-of-water excursions got started there were aquatic creatures. (Of course! There are fish today, right?) But at a given geological time, e.g., 365 million years ago, some creatures that were more aquatic have been discovered, such as Acanthostega, as well as some less aquatic ones, such as Ichthyostega. All of them are transitional, and not true land-dwellers yet. They cannot even be called amphibians, they’re between fish and amphibians.
If we want to meet some true amphibians, we have to move several dozen million years forward in time. Compare the above with Eryops, shown below.
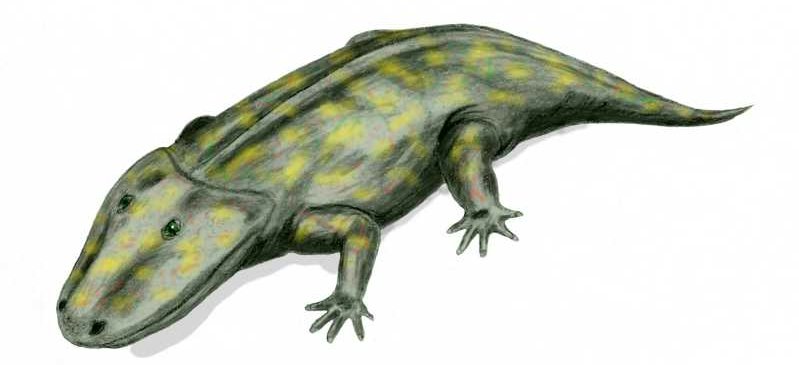
Eryops megacephalus, an early Permian (~295 M. y. ago) amphibian (Credit: Arthur Weasley, GNU Free Documentation licence)
The Eryops (meaning “drawn-out face”), shown above, was a true amphibian that lived around 295 million years ago. Notice its tail which lacks the finned end of the previous tetrapodomorphs, and its feet, which are strong, not webbed, and with five fingers. By the way, the number of fingers is very telling about the animal’s way of life: generally speaking, a larger number of fingers/toes shows a more aquatic way of living, and less reliance on coming out of the water. Look at the following figure:
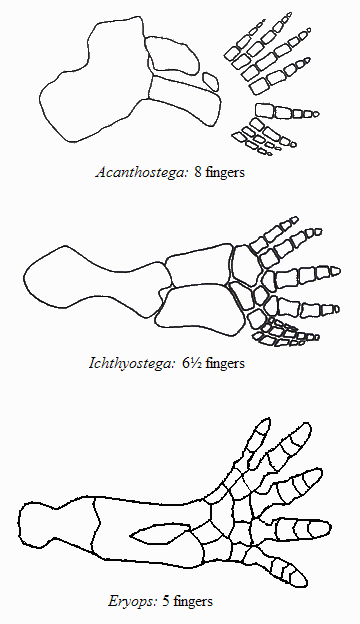
Comparison of limb structure of the mentioned fossils (after Cowen and Jarvik)
We see that Acanthostega, which lived a more aquatic life among the above-mentioned creatures, had 8 fingers (or toes); Ichthyostega, more often needing to support its weight on its libs, had 6 “good” fingers and one vestigial one; its foot perhaps was on its way to become 6-toed; and Eryops, a true amphibian, had five fingers — notice also how much stronger they are. This reduction in the number of fingers is only natural: an aquatic vertebrate uses its limbs as oars, so the fingers need to be thin and many, to make a flexible webbed oar; whereas a terrestrial vertebrate needs strong fingers/toes to support its weight on them, and being stronger they don’t have enough space at the end of the foot, so they are reduced in number.
After the end of the Devonian period, and during the period called Carboniferous (360 – 300 M. y. ago), we amphibians became very many. But really-really very many. Later, however, by the middle of the next period (called Permian, 300 – 250 M. y. ago), our dominance was dealt a severe blow: the reptiles came, including your mammalian ancestors, and essentially displaced us, pushing us out of every niche that we had occupied, save for a few very specialized ones.
So what happened to us amphibians? Why is our once-glorious class now represented by tiny animals, such as mainly us frogs, and the occasional salamander and newt?

A modern amphibian: the tiny Emperor Newt (Tylototriton shanjing) © (*). Compare its body structure with Eryops.
Oh, it’s a sad story. I better let the human who lent me the space in this page to explain why we amphibians lost our dominance and appear so inconspicuous today.
That was the froggy’s story, recounting how our common ancestors (common between us humans and the frogs) gradually came out of the water during those fateful millions of years, in the late Devonian period: gradually, and yet “abruptly”. Putting now the above-mentioned fossils onto the colored strip that was shown earlier, we obtain roughly the following diagram:
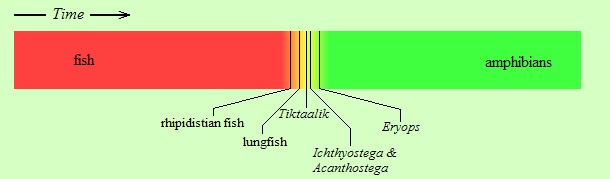
So we see that in the case of the fish-to-amphibian transition we were lucky enough to have found just the right kind of fossils that document precisely that transition. Naturally, the search is not over, so the collection keeps being enriched, decade after decade.
But the documentation of this transition rests on the tireless efforts of paleontologists who go in search of fossils and dig the rocks, often in hostile, forbidding places, such as northern Canada and Greenland. In other cases, scientists have not been so lucky, or there hasn’t been so much effort exerted yet. After all, the geological past time is vast, the evolutionary tree enormous, and the number of transitions from species to species overwhelming. Still, creationists go and look between the vertical black lines in the figure above, demanding fossils even there (forgetting that the gaps used to be wider in the past, and that creationists were demanding fossils in those gaps, and the fossils were found!). Just how narrow must the gaps be before creationists admit that there is a continuity? (And how narrow must the minds be to keep seeing gaps instead of the continuity?)
Similarly compelling transitional fossils have been discovered in the reptile-to-bird transition, and in the common-ape-to-intelligent-ape (human) transition. But, as I said, no matter how many and compelling the findings, creationists always don the air of experts and pontificate: this is fish, the other is amphibian; this is ape, that other one is human. For them, the world is — alas — too simple: either black or white.
The above colored strip might give to the reader the impression that evolution proceeds linearly: from fish to amphibians, then to reptiles, and so on. Nothing can be further from the truth: as I already mentioned, evolution proceeds in a tree-like fashion, not in a chain-like fashion. For example, even after the late Devonian period when the first amphibians and later the reptiles appeared, fish continued to evolve in the water, and new fish-forms appeared that were not present back then. Likewise with amphibians: there were no frogs among the first amphibians; frogs are late comers in the amphibian class, they appeared after the emergence and dominance of the reptiles.
To visualize the idea that evolution proceeds in a tree-like fashion and not linearly, and also the idea of adaptive radiation, consider the following animated drawing. As before, the red part represents aquatic environments (sea, rivers, lakes), and the green represents terrestrial environments (land). The first vertebrates appear first as a short blue line on the left, in the water, and they start radiating, which means, branching quickly and making up many related but different species. Each of these species occupies a niche. Then these species branch again, and many more species are created, until all the available niches within the aquatic (red) environment are occupied. Now take a look and see what happens after all the niches in the red area are occupied in the animated figure below:

Between the aquatic and terrestrial area there is a black division with some “aisles”. These aisles represent some specialized niches that exist “between” water and land: niches such as those that Tiktaalik, Ichthyostega, and the rest of the extinct transitional creatures occupied. I drew them as narrow aisles to reflect the observation that only very specific conditions in evolution will lead through them. After the middle aisle — which is the only one that leads vertebrates from water to land in this drawing — is occupied, species after species, vertebrates (now transformed to amphibians) find themselves in an unoccupied territory (green), a terra incognita, a land with a large number of environments where no other large-bodied animals exist yet. (There are only insects and plants on land at that time.) But, as is well known, “nature abhors a vacuum”, and so adaptive radiation starts once more, this time on land, with amphibians and soon the rest of the terrestrial vertebrates (reptiles, mammals, birds) occupying every terrestrial nook and cranny. Notice that at some point niches that lead back to the water are explored (see the top-most aisle).(*) These are supposed to be mammals like the ancestors of whales and dolphins, which are related to today’s even-toed mammals (the Artiodactyla), such as cows, hippos, and pigs. At a later point their decendants (whales and dolphins) really ventured permanently into the water, an event represented by the late branch that goes well into the red area. But whales and dolphins do not radiate again into the water — why? Because most niches are already occupied there, by the fish! If the fish were miraculously wiped out from our planet, then whales and dolphins would radiate adaptively in the water, making it inhabited and teeming with life once again, this time by aquatic mammals. Other aisles lead also back toward the water but not quite (think of seals, sea otters, the walrus, water snakes, crocodiles, and the like).
Notice that there is something not shown in this animation: species do not only appear, they are also extinct, and no previously drawn line is ever deleted in the animated figure. Well, I couldn’t work endlessly to make this animation even more realistic. Besides, remember to interpret each branching point not as a species, but as a niche in that figure. So, even though species are extinct, niches are somewhat more persistent in time (though not eternal).
Why isn’t the land occupied again and again by fish? Why don’t the likes of Tiktaalik appear again?
Part of the answer is given by the diagram above: niches that would lead from water to land are already occupied, and when a niche is occupied you can’t very easily push its inhabitant out (drive it to extinction), and occupy it yourself. Even if this happens once, then your descendant species would have to push again another occupant out of its specialized niche, along the evolutionary “path” that leads to land. (That’s an abstract path, of course, not a physical one.) So, a series of unlikely events would have to happen, which makes the probability exceedingly small. Still, it is not impossible that it can happen. Today, for example, there are species of eel (which are bony fish) that slither on moist land (e.g., grass) and move from pond to pond. It is not impossible that after several million years their descendant species will be even more land-going, competing in niches that today are occupied by snakes. One never knows what evolution has in stock for the future.
The adaptive radiation that occurred when vertebrates came out of water and occupied the land was not an exceptional event. Such events have happened all the time. Another such grand-scale adaptive radiation event was when some reptiles (Archeopteryx and its relatives) developed feather-like structures, perhaps initially for gliding from tree to tree, as some modern squirrels do, or for leaping onto trees for refuge — the exact environmental pressure is still unknown — thus becoming better and better at gliding, and then flying, eventually conquering the high air, which provides plenty of new and unoccupied niches. Thus we got the birds. Darwin himself observed another, smaller-scale result of adaptive radiation: the numerous species of finches on the Galápagos islands: finches that arrived there found no other birds of their size on the islands, and so they discovered their own terra incognita (of niches), radiating into a large number of finches. (That was a crucial observation for Darwin while he was developing the theory of evolution.) There have been other, more spectacular events of adaptive radiation (at least from our human point of view), ones that most likely will never happen again (at that scale — unless all multicellular creatures are wiped out). One is the radiation during the Ediacaran period, (~630 to 540 M. y. ago) when multicellular creatures emerged; and the other is the event of the Cambrian explosion, when some of the Ediacaran animals evolved and found the entire Earth unoccupied by beings of their size. At that time (~530 M. y. ago, during the Cambrian period) the basic body architectures for animals were designed. Many of them perished, and the ones that remain today form the various phyla of animals, i.e., the most general groups in the animal kingdom, such as the mollusks (snails, bivalves, octopuses, and such), the arthropods (insects, spiders, crabs, and such), the chordates (vertebrates and a couple more groups — that’s the phylum where we humans are), and several more phyla.
Now, the froggy appeared a bit disappointed in the end while describing how amphibians lost their dominant role on land, right? What could the froggy mean? There is no small number of amphibian species today. Here is approximately how many species exist in each of the vertebrate classes:
(The jawless fish is an even earlier class of fish than the cartilaginous ones.) So, excluding the bony fish, of which there are really many, amphibians do not really fare too badly in terms of number of species; they’re clearly more than mammals. (And “reptiles” is not just one evolutionary branch, it’s a grab-bag for “terrestrial vertebrate that’s neither amphibian, nor mammal, nor bird”.) What the froggy meant was that the amphibians appeared to be the lords on land (of some land, close to water) at one time, but were then forced by the rest of the land-going vertebrates (reptiles and mammals) to retreat to specialized niches. Initially amphibians had the forms of some of today’s reptiles, such as the crocodiles, but later such forms went extinct by similar reptiles. Today amphibians are mostly tiny animals — most of them are frogs and toads — and they survive at the “edge” of environments, where freshwater meets land. In that sense they are “pushed into a corner”, and represent “left overs” of a once thriving category of creatures.
There are many other examples of such “left over” categories of beings, and their study (or mere knowledge of them) gives us a deeper understanding of how evolution proceeds. There are left-over mammals (the monotremes, 5 species only) that lay eggs; left-over birds (the Struthioniformes: ostrich, emu, and a few more, 10 in total), that are large-bodied and don’t fly; left-over reptiles, the tuataras (only two kinds) that possess a third eye; left-over fish, which we met already, such as the lobe-finned ones (coelacanths, lungfish: 8 species); left-over insects, such as the silverfish, which don’t fly; and even pre-insects, such as the centipedes and millipedes; and so on. You name a group of living beings, and you can find a left-over group among them, almost always.
“Left overs” are really hard to justify as the outcome of the work of an intelligent designer. Such a designer would appear as if he was changing his mind all the time. For example, the designer first made some six-legged wingless insects, and then thought, “Oh!... wait a minute: if these things could grow wings they would make a helluva — oops! sorry — diverse taxon [biological group].” Then he put wings on insects, and the winged thingies radiated (to over a million identified species today), displacing the original wingless forms, of which only rare kinds like silverfish exist today. Is this really intelligent design?
But the fascinating lesson of the “left overs” of evolution does not belong to this page, which is about religious dishonesty and the distortion of the theory of evolution by creationists. Creationists do not even dare to mention concepts like the “left overs” — if they are aware of them at all — because they want notions that they think are hard for biologists to explain, not others that are hard for them, because they can’t be reconciled with the notion of intelligence in design. So I’ll postpone developing the left-overs topic, relegating it to a different category of subjects that refute creationism.
Recall that this page (as well as the previous one) originated as part of a series of pages that answer the supposed evolutionary conundrums raised by the folks at Multimedia Apologetics. Two more such “conundrums” were raised by them, shown below:
3. The theory of evolution violates the second law of thermodynamics
4. If life arose through evolution, why can’t scientists create life in the laboratory?
Click on the linked topic if you are interested in knowing what the answer is.
| Footnotes: (clicking on the caret
(^) on the left of the footnote brings back to the text) (^) In the fifth edition of The Origin of Species Darwin wrote: “the periods during which species have undergone modification, though long as measured in years, have probably been short in comparison with the periods during which they retain the same form.” (^) I shied from writing Lamarck’s full name in the main text. Here it is: Chevalier de Jean Baptiste Pierre Antoine de Monet Lamarck. (^) However, this is not very far from the modern understanding of one of the mechanisms of evolution: catastrophic geological events causing extinctions can result in significant and “sudden” evolutionary “innovation”, and in adaptive radiation, a term that will be explained later. (^) Of course another, much more common alternative, is that the environmental pressure results in the extinction of the species. (^) Note that some biologists such as Richard Dawkins have claimed that the idea of constant gradual change is an extreme case and a caricature of evolution, and that the speed along which evolution proceeds is variable. Dawkins considers the idea of “punctuated equilibrium”, to be discussed immediately, as just another extreme case. (^) The American biologist G. Ledyard Stebbins has calculated that an animal species could grow from mouse-sized to elephant-sized within 60,000 years (in R. Dawkins, The Blind Watchmaker, 1996 edition, W. W. Norton, p. 242). (^) Punctuated equilibrium was clearly stated in a 1972 landmark paper by paleontologists Niles Eldredge and Stephen Jay Gould, but was hinted at by Ernst Mayr in 1954, and even by Charles Darwin (see footnote, above). It has been criticized by Dawkins (among others), who, in chapter 9 of The Blind Watchmaker (see reference in previous footnote), claims that sudden appearances of species in the fossil record are most likely due to migrational events: species evolved elsewhere, then migrated, and fossilized where found. Though this is possible, and is probably the explanation for a number of cases, it sounds like an extremist position if invoked in all cases: it does not explain why intermediate species are usually not found even elsewhere (but see notable exceptions in the rest of this page). (^) Did you notice that at some point the top-most aisle is extended and opens up a pathway that leads from land back to water? With this I wanted to underline the observation that niches are not fixed and eternal; they are constantly shifting, and often it is living creatures themselves that create new niches for other living beings that come and occupy them. |
Comments, opinions, or suggestions? Email to the author
Back to the Topics against Creationism
Created: March 29, 2008
© Copyright notice: All pictures of living animals in this page belong to and are copyrighted by the author. The author is not interested in making this material appear in print. However, any attempt to copy its content in articles that appear either in print, or on the web, or other media of information without an explicit reference to their source, i.e., this web page, will be considered an unethincal act of plagiarism (at best) and a theft of intellectual property (at worst). Please note that intellectual property is automatically protected by law in almost all countries of the world.